Subject: Re: mechanical cochlear model From: David Mountain <dcm@xxxxxxxx> Date: Tue, 9 Mar 2010 20:59:19 -0500 List-Archive:<http://lists.mcgill.ca/scripts/wa.exe?LIST=AUDITORY>--00151747832e7145c9048168a51d Content-Type: text/plain; charset=ISO-8859-1 Many theories have been advanced to explain the details of the peak region of the cochlear response but perhaps we can try to agree on some fundamental principals. I would argue that a traveling wave, involving transverse motion of the basilar membrane, passively propagates acoustic energy to the peak region. This passive propagation behaves in a manner described by Peterson and Bogert (1950). When this energy reaches the beginning of the peak region, some process, usually called the "cochlear amplifier" and involving outer hair cells, takes over and shapes the peak response. We may question whether the cochlear amplifier results from somatic motility or from hair bundle motility and we may debate questions of how outer hair cell forces get coupled back into basilar membrane and inner hair cell responses but does anyone question whether the classical traveling wave plays a role in at least passive propagation of acoustic energy to the best place? On Mon, Mar 8, 2010 at 9:25 PM, Lloyd Watts <lwatts@xxxxxxxx> wrote: > Rhode's 1971 data showed about 3-4 cycles of phase accumulation to the best > place, independent of amplitude. I believe that is strong evidence of a > traveling wave that takes 3-4 cycles to arrive, and I believe from > discussions directly with Bill in 2000-2003 that this is also how he > interpreted his measurements. > > Regarding the stiffness change not being sufficient to explain 3 orders of > magnitude of frequency, I believe that argument is based on the assumption > of constant membrane mass (what I called in my thesis, the constant-mass > scaling assumption, which does indeed imply 6 orders of magnitude of change > of stiffness from base to apex, and the damping decreases by 3 orders of > magnitude). But the widening of the basilar membrane is more consistent > with an increasing-mass assumption, in which the stiffness decreases, the > mass increases, and the damping remains constant. Neither is probably > exactly correct, but it shows that having all the change in the stiffness is > not necessary, and the widening of the basilar membrane is consistent with > both a mass increase and a stiffness decrease. > > The original discussion began around a tutorial objective - how to explain > the basic idea of cochlea operation to a beginner, with a mechanical model > that could demonstrate the wave propagation in real time. My preferred way > is to describe the 2D model (much as Alain eloquently did, although I > usually use a picture), and show the following animated figure for the 2D > model, which is sufficient to illustrate the main behaviors of the passive > model: > > http://www.lloydwatts.com/1600mov2.gif > > from my thesis (page 19, Figure 2.8), which shows the long-wave, > short-wave, and cutoff behaviours, all with smooth transitions from one to > the other, showing both membrane displacement and fluid pressure in the 2D > duct. And then to explain that the 2D passive model can explain the passive > behavior at high sound levels - the active processes of the outer hair cells > are necessary to explain the extra gain and sharpness found at low sound > levels, as in Rhode's 1971 data. Of course, the fluid pressure figure could > be flipped over and inverted to illustrate more explicitly the behavior of > the upper and lower ducts. > > Finally, I note that the animation above is really just a visualization of > one of the curves from Steele and Taber, 1979, "Comparison of WKB and finite > difference calculations for a two-dimensional cochlear model", JASA 65, > 1001-1006. I replotted those curves here: > http://www.lloydwatts.com/cochlea.shtml > > along with my mode-coupling correction that fixes up the incorrect behavior > in the cut-off region - a second wave mode is needed to explain the change > of slope and occasional notch which is observed after the best place. I > note that the combination of the plausible 2D simulation model that can be > animated, with the Liouville-Green (or WKB) analytic model is a powerful > combination for both personal understanding and for tutorial explanation to > beginners. > > The mode-coupling solution that reconciles the simulation model and the LG > (or WKB) model in all regions, is published in my 1992 thesis, available > online: > http://www.lloydwatts.com/thesis.html > and also was published in > L. Watts, "The mode-coupling Liouville-Green approximation for a > two-dimensional cochlear model", Nov. 2000, JASA 108, pp. 2266-2271. > > Thank you, > Lloyd Watts > > > -----Original Message----- > From: AUDITORY - Research in Auditory Perception [mailto: > AUDITORY@xxxxxxxx On Behalf Of Matt Flax > Sent: Monday, March 08, 2010 2:16 PM > To: AUDITORY@xxxxxxxx > Subject: Re: [AUDITORY] mechanical cochlear model > > The concept that the passive inner ear mechanism does not conduct energy > as a fast pressure wave (with long wavelength) may be true - however not > everyone agrees on this point. I believe that you are talking about the > coupling between the outer ear and the inner ear below - and thus the > passive action of the hearing system. > > The concept that the active inner ear mechanism is an active travelling > wave seems a little impossible (from what people have discussed > previously - experimental evidence and theory) for any location other > then the base of the inner ear. I stand on record as the first for > claiming that the base is dominated by the active travelling wave and > the mid/apex is dominated by the active long pressure wave (my > compression wave cochlear-amplifier definition). > > Matt > > On Mon, 2010-03-08 at 07:52 -0800, Richard F. Lyon wrote: > > Alain, your "simple account" was "reasonably correct" whether you > > mentioned the traveling wave or not, so I agreed with you. > > > > I admit I don't see why you don't explicitly identify that account > > with traveling waves. Sinusoids in such a system are locally > > described as cos(ometa*t - k*x), with some varying amplitude, where > > the relations between omega and k locally obey the same kind of > > dispersion relation (more or less) as gravity waves on water. > > > > The real issue seems to be about where the energy is, and how it > > propagates and gets detected. The fast pressure wave is another > > physical mode that really exists, but its wavelengths are all very > > large compared to the cochlea, so the pressure due to this mode can > > be viewed as equal throughout the cochlea; pushing on the stapes > > immediately pushes out at the round window. But this mode doesn't > > carry much energy, as the middle ear leverage isn't nearly enough to > > couple efficiently to it; there's also no efficient way to get energy > > back out of this mode into a place and a form for which detectors are > > known; the fast compression wave therefore carries pressure, but not > > much energy. The traveling wave mode, on the other hand, involves > > much larger fluid displacements at the same pressures; it's a > > differential mode between the scalae, propagated by the spring of the > > BM instead of by fluid compression. And the energy converges on the > > BM as the wave slows down and the wavelength gets short, focusing the > > energy into a small layer with large displacements at the BM. There, > > the hair cells, which evolved to detect fluid motion via cilia > > bending, are well positioned to respond. > > > > A wave has three kinds of delays: phase, group, and wave-front. In > > typical models, the wave-front delay is essentially zero, and the > > response latency can be made to approach zero as the level gets very > > high. The group delay depends on the damping, including negative > > damping effects, and so varies a lot with level. The phase delay is > > in between, around a cycle and half, and pretty stable. Pretty much > > everything about it is as Lighthill described, in terms of energy > > flow, except that there's not much BM mass so it's not really > > significantly resonant, except for very high frequencies very near > > the base where the accelerations are very high. And except for some > > positive feedback from outer hair cells that modifies the dispersion > > relation, providing active gain instead of loss over some range of > > wavelengths. > > > > If it walks like a wave, and quacks like a wave, and transports > > energy like a wave, why not call it a wave? > > > > Dick > -- David C. Mountain, Ph.D. Professor of Biomedical Engineering Boston University 44 Cummington St. Boston, MA 02215 Email: dcm@xxxxxxxx Website: http://www.bu.edu/hrc/research/laboratories/auditory-biophysics/ Phone: (617) 353-4343 FAX: (617) 353-6766 Office: ERB 413 --00151747832e7145c9048168a51d Content-Type: text/html; charset=ISO-8859-1 Content-Transfer-Encoding: quoted-printable Many theories have been advanced to explain the details of the peak region = of the cochlear response but perhaps we can try to agree on some fundamenta= l principals. =A0I would argue that a traveling wave, involving transverse = motion of the basilar membrane, passively propagates acoustic energy to the= peak region. =A0This passive propagation behaves in a manner described by = Peterson and Bogert (1950). =A0When this energy reaches the beginning of th= e peak region, some process, usually called the "cochlear amplifier&qu= ot; and involving outer hair cells, takes over and shapes the peak response= . =A0We may question whether the cochlear amplifier results from somatic mo= tility or from hair bundle motility and we may debate questions of how oute= r hair cell forces get coupled back into basilar membrane and inner hair ce= ll responses but does anyone question whether the classical traveling wave = plays a role in at least passive propagation of acoustic energy to the best= place?<div> <br><br><div class=3D"gmail_quote">On Mon, Mar 8, 2010 at 9:25 PM, Lloyd Wa= tts <span dir=3D"ltr"><<a href=3D"mailto:lwatts@xxxxxxxx">lwatts@xxxxxxxx= ience.com</a>></span> wrote:<br><blockquote class=3D"gmail_quote" style= =3D"margin:0 0 0 .8ex;border-left:1px #ccc solid;padding-left:1ex;"> Rhode's 1971 data showed about 3-4 cycles of phase accumulation to the = best place, independent of amplitude. =A0I believe that is strong evidence = of a traveling wave that takes 3-4 cycles to arrive, and I believe from dis= cussions directly with Bill in 2000-2003 that this is also how he interpret= ed his measurements.<br> <br> Regarding the stiffness change not being sufficient to explain 3 orders of = magnitude of frequency, I believe that argument is based on the assumption = of constant membrane mass (what I called in my thesis, the constant-mass sc= aling assumption, which does indeed imply 6 orders of magnitude of change o= f stiffness from base to apex, and the damping decreases by 3 orders of mag= nitude). =A0But the widening of the basilar membrane is more consistent wit= h an increasing-mass assumption, in which the stiffness decreases, the mass= increases, and the damping remains constant. =A0Neither is probably exactl= y correct, but it shows that having all the change in the stiffness is not = necessary, and the widening of the basilar membrane is consistent with both= a mass increase and a stiffness decrease.<br> <br> The original discussion began around a tutorial objective - how to explain = the basic idea of cochlea operation to a beginner, with a mechanical model = that could demonstrate the wave propagation in real time. =A0My preferred w= ay is to describe the 2D model (much as Alain eloquently did, although I us= ually use a picture), and show the following animated figure for the 2D mod= el, which is sufficient to illustrate the main behaviors of the passive mod= el:<br> <br> <a href=3D"http://www.lloydwatts.com/1600mov2.gif" target=3D"_blank">http:/= /www.lloydwatts.com/1600mov2.gif</a><br> <br> from my thesis (page 19, Figure 2.8), which shows the long-wave, short-wave= , and cutoff behaviours, all with smooth transitions from one to the other,= showing both membrane displacement and fluid pressure in the 2D duct. =A0A= nd then to explain that the 2D passive model can explain the passive behavi= or at high sound levels - the active processes of the outer hair cells are = necessary to explain the extra gain and sharpness found at low sound levels= , as in Rhode's 1971 data. =A0Of course, the fluid pressure figure coul= d be flipped over and inverted to illustrate more explicitly the behavior o= f the upper and lower ducts.<br> <br> Finally, I note that the animation above is really just a visualization of = one of the curves from Steele and Taber, 1979, "Comparison of WKB and = finite difference calculations for a two-dimensional cochlear model", = JASA 65, 1001-1006. =A0I replotted those curves here:<br> <a href=3D"http://www.lloydwatts.com/cochlea.shtml" target=3D"_blank">http:= //www.lloydwatts.com/cochlea.shtml</a><br> <br> along with my mode-coupling correction that fixes up the incorrect behavior= in the cut-off region - a second wave mode is needed to explain the change= of slope and occasional notch which is observed after the best place. =A0I= note that the combination of the plausible 2D simulation model that can be= animated, with the Liouville-Green (or WKB) analytic model is a powerful c= ombination for both personal understanding and for tutorial explanation to = beginners.<br> <br> The mode-coupling solution that reconciles the simulation model and the LG = (or WKB) model in all regions, is published in my 1992 thesis, available on= line:<br> <a href=3D"http://www.lloydwatts.com/thesis.html" target=3D"_blank">http://= www.lloydwatts.com/thesis.html</a><br> and also was published in<br> L. Watts, "The mode-coupling Liouville-Green approximation for a two-d= imensional cochlear model", Nov. 2000, JASA 108, pp. 2266-2271.<br> <br> Thank you,<br> <font color=3D"#888888">Lloyd Watts<br> </font><div class=3D"im"><br> <br> -----Original Message-----<br> From: AUDITORY - Research in Auditory Perception [mailto:<a href=3D"mailto:= AUDITORY@xxxxxxxx">AUDITORY@xxxxxxxx</a>] On Behalf Of Matt F= lax<br> Sent: Monday, March 08, 2010 2:16 PM<br> To: <a href=3D"mailto:AUDITORY@xxxxxxxx">AUDITORY@xxxxxxxx</a= ><br> Subject: Re: [AUDITORY] mechanical cochlear model<br> <br> </div><div><div></div><div class=3D"h5">The concept that the passive inner = ear mechanism does not conduct energy<br> as a fast pressure wave (with long wavelength) may be true - however not<br= > everyone agrees on this point. I believe that you are talking about the<br> coupling between the outer ear and the inner ear below - and thus the<br> passive action of the hearing system.<br> <br> The concept that the active inner ear mechanism is an active travelling<br> wave seems a little impossible (from what people have discussed<br> previously - experimental evidence and theory) for any location other<br> then the base of the inner ear. I stand on record as the first for<br> claiming that the base is dominated by the active travelling wave and<br> the mid/apex is dominated by the active long pressure wave (my<br> compression wave cochlear-amplifier definition).<br> <br> Matt<br> <br> On Mon, 2010-03-08 at 07:52 -0800, Richard F. Lyon wrote:<br> > Alain, your "simple account" was "reasonably correct&qu= ot; whether you<br> > mentioned the traveling wave or not, so I agreed with you.<br> ><br> > I admit I don't see why you don't explicitly identify that acc= ount<br> > with traveling waves. =A0Sinusoids in such a system are locally<br> > described as cos(ometa*t - k*x), with some varying amplitude, where<br= > > the relations between omega and k locally obey the same kind of<br> > dispersion relation (more or less) as gravity waves on water.<br> ><br> > The real issue seems to be about where the energy is, and how it<br> > propagates and gets detected. =A0The fast pressure wave is another<br> > physical mode that really exists, but its wavelengths are all very<br> > large compared to the cochlea, so the pressure due to this mode can<br= > > be viewed as equal throughout the cochlea; pushing on the stapes<br> > immediately pushes out at the round window. =A0But this mode doesn'= ;t<br> > carry much energy, as the middle ear leverage isn't nearly enough = to<br> > couple efficiently to it; there's also no efficient way to get ene= rgy<br> > back out of this mode into a place and a form for which detectors are<= br> > known; the fast compression wave therefore carries pressure, but not<b= r> > much energy. =A0The traveling wave mode, on the other hand, involves<b= r> > much larger fluid displacements at the same pressures; it's a<br> > differential mode between the scalae, propagated by the spring of the<= br> > BM instead of by fluid compression. =A0And the energy converges on the= <br> > BM as the wave slows down and the wavelength gets short, focusing the<= br> > energy into a small layer with large displacements at the BM. =A0There= ,<br> > the hair cells, which evolved to detect fluid motion via cilia<br> > bending, are well positioned to respond.<br> ><br> > A wave has three kinds of delays: =A0phase, group, and wave-front. =A0= In<br> > typical models, the wave-front delay is essentially zero, and the<br> > response latency can be made to approach zero as the level gets very<b= r> > high. =A0The group delay depends on the damping, including negative<br= > > damping effects, and so varies a lot with level. =A0The phase delay is= <br> > in between, around a cycle and half, and pretty stable. =A0Pretty much= <br> > everything about it is as Lighthill described, in terms of energy<br> > flow, except that there's not much BM mass so it's not really<= br> > significantly resonant, except for very high frequencies very near<br> > the base where the accelerations are very high. =A0And except for some= <br> > positive feedback from outer hair cells that modifies the dispersion<b= r> > relation, providing active gain instead of loss over some range of<br> > wavelengths.<br> ><br> > If it walks like a wave, and quacks like a wave, and transports<br> > energy like a wave, why not call it a wave?<br> ><br> > Dick<br> </div></div></blockquote></div><br><br clear=3D"all"><br>-- <br><br>David C= . Mountain, Ph.D.<br>Professor of Biomedical Engineering<br><br>Boston Univ= ersity<br>44 Cummington St.<br>Boston, MA 02215<br><br>Email: =A0 <a href= =3D"mailto:dcm@xxxxxxxx">dcm@xxxxxxxx</a><br> Website: <a href=3D"http://www.bu.edu/hrc/research/laboratories/auditory-bi= ophysics/">http://www.bu.edu/hrc/research/laboratories/auditory-biophysics/= </a><br>Phone: =A0 (617) 353-4343<br>FAX: =A0 =A0 (617) 353-6766<br>Office:= =A0ERB 413<br> <br> </div> --00151747832e7145c9048168a51d--
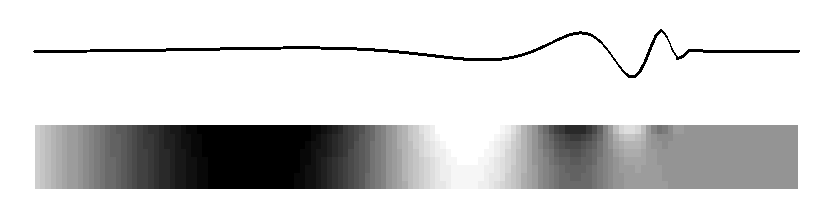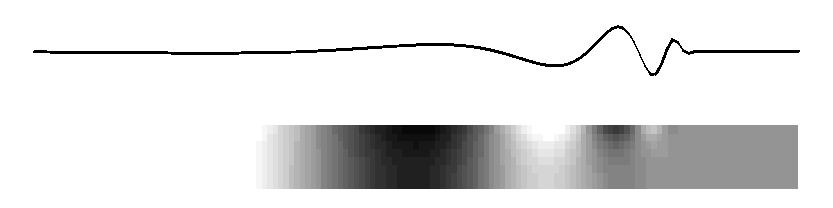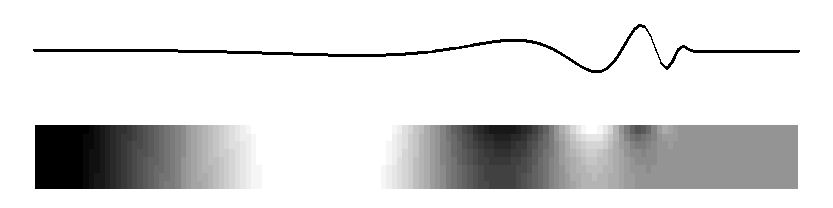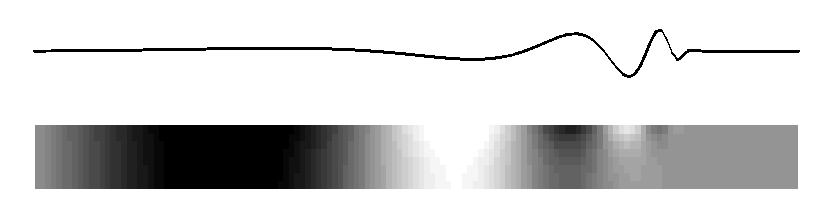{kind=link}